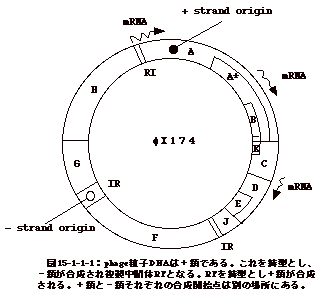
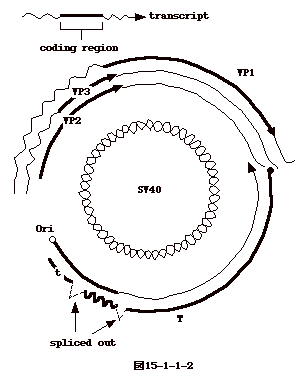
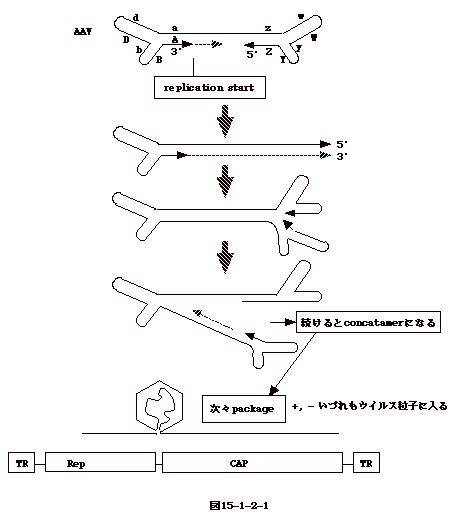
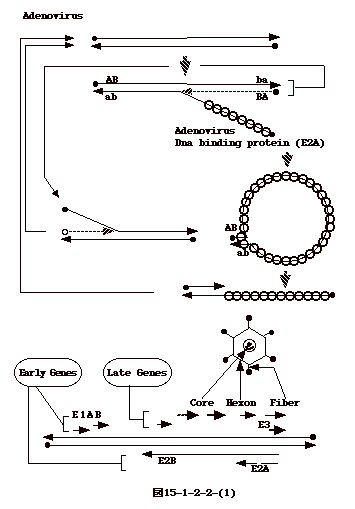
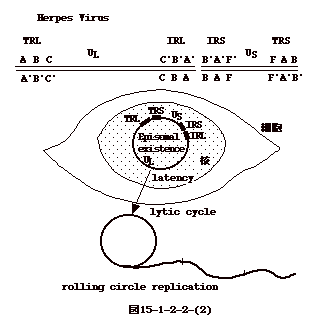
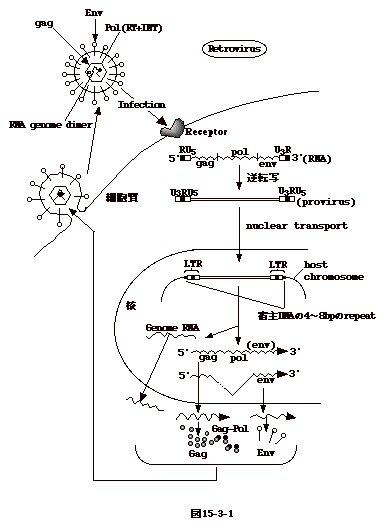
15-3-1:レトロウイルス
Retrovirusは次の3群に分けられる。
Oncornavirus: マウスやトリの白血病ウイルス、 HTLV-1
Lentivirus : HIV、SIV
Spumavirus : foamy virus
ウイルス粒子はゲノムRNAの二量体(dimer)を含む。感染すると細胞質でDNAに読み取られ、両端にLTRと云う構造を持つプロウイルスとなる。プロウイルスはウイルス粒子を構成するPol蛋白と共に核に移行し(恐らくGag或いはGag-Pol蛋白と一緒に)、細胞の染色体DNAに組み込まれる。組替え部位を調べると、組み込まれたプロウイルスの両端に数塩基の繰り返し配列が見られる。これは、トランスポソンに特徴的な現象である。組み込まれたDNAから転写されたRNAは再びウイルス粒子に取り込まれる。
ウイルス蛋白としては、ウイルス粒子のコアを形成するGag蛋白、エンベロープ蛋白(Env)、逆転写(RT)と組み込み(INT)酵素の活性を持つPol蛋白がある。Pol(Gag-Pol融合蛋白として翻訳)及びGagはゲノムサイズのRNAから、Envはスプライスによりgag-pol部分が除かれたRNAから翻訳される(図15-3-1)。
以上は、最も単純な構造のOncornavirusの場合であるが、他のレトロウイルスもほぼ同様な機構で複製する。但し、LentivirusやSpumavirusでは、ウイルス遺伝子の発現を調節する遺伝子が3'末の方に存在する点がOncornavirusと異なる。又、前者のタイプのウイルスは生殖系列細胞に組み込まれ、内在性ウイルスとなり得るが(HTLV-1を除く)、他のウイルスではその様な事は報告されていない。
レトロウイルスの中では、エイズの原因であるHIV、成人T細胞性白血病の原因であるHTLV-1が重要である。
特にHIVは、世界的な広がりを見せ、1996年の段階で、2千3百万の感染者がおり、既に6百万が死亡している。感染者の9割は途上国にいる。
わが国でも平成9年辺りから献血者にHIV陽性者が増え始め、日本の社会に広がりつつある事が示唆されている。日本のHIV感染は、1983年当初、HIV汚染血液製剤による血友病患者が大半を占めていたが、次第に性感染、特に異性間性交渉によるものが増加している。
2002年現在では20〜30台の男性同性間感染が目立って多くなっている。
HIVは、血液又は性交渉により感染する。感染により、免疫の中枢とも云えるCD4陽性ヘルパーT細胞を破壊する。血液中のCD4細胞の減少に伴い免疫低下による真菌症、カリニ肺炎、細菌やウイルスの感染、カポジ肉腫と呼ばれる血管腫などのエイズ(Acquired
Immunodeficiency Syndrome, AIDS)の症状が現れる。経過には個人差があるが、感染初期のHIVウイルス量が多くCD4値の低い者程経過が早く死に至る。面白い事に感染してもエイズ発症をしない長期生存者が存在する。マウスの白血病ウイルスの実験で良く分かっているが、レトロウイルスの感染や発病を支配する遺伝子が存在し、HIVにもそのような現象がある、と云う事である。
HIVの感染はヘルパーT細胞のCD4分子をウイルス外皮蛋白が認識し結合する事により開始するが、CD4分子のみでは十分でなくcoreceptorが必要である事が分かって来た。HIVウイルスにはマクロファージに感染するものとT細胞に感染するものとがあるが、その違いはcoreceptorが異なる為のようである。
HIV感染に対しては、ウイルスの持つ逆転写酵素やプロテアーゼに対する阻害剤で治療効果を挙げられるようになった。但し、薬剤耐性ウイルスの出現などにより治療が効を奏さない場合がある
HIVに関しては、Levyの HIV and the Pathogenesis of AIDS, 2nd Edition, ASM Press
(1998)が示唆に富む。レトロウイルス一般については、Coffin, Hughes,
Varmusの Retroviruses. Cold Spring Harbor Laboratory Press (1997)が面白い。
米国ではHIV汚染血液製剤により、日本以上の犠牲者が出た。この事に関し、米国では調査委員会が設置され、これが対処した。その調査報告書、 L.B.
Leventon, H.C. Sox and M.A. Stoto(eds.). HIV and the Blood Supply - Analysis
of Crisis Decisionmaking Institute of Medicine, National Academy Press, 1995
ISBN 0-309-05329-3
が出ている。この調査は科学的側面、行政のあり方の解析など学ぶ点が多いので一読を勧める。特に、このような調査の危険と利益について、「利益とは次の機会への教訓であり、危険は現在の知識により当時得られた知識の限界の中で意志決定をせざるを得なかった人々への不当な批判である」としている。
HIV汚染血液製剤の問題については、幾つか、冷静に、考える必要がある。幾つかのポイントを以下に示す。
- HIVの現在認識されているような危険性が認められるようになったのはどの時点か。
- エイズがウイルスによると云う説が受け入れられたのは何年位の頃か。
-
日本で1983年の段階でいわゆる製剤からクリオプレシピテート(cryoprecipitate)に変えるべきであったと云う議論があったが、クリオとはどういう利点と欠点があったか。
- 何時完成した加熱製剤が出来上がったのか(1988年に加熱製剤のみを投与された患者でエイズ発症が報告されている)。
-
医薬品の市場引き上げの制度はどうなっているのか。米国と日本の違いはどうか。何故、米国業者は1983年と云う早い段階で汚染血液を原材料とした製剤の市場引き上げをしたのに、何故日本では遅れたのか。
- 厚生省の仕組みには、その担当者の人的構成を含め問題がなかったか。今、現在、それは解決されているのか。
これらへの、筆者の考えは、「公衆衛生と感染症」(対象としての人間:岩波講座科学/技術と人間6、126-156頁)に概略を示してある。
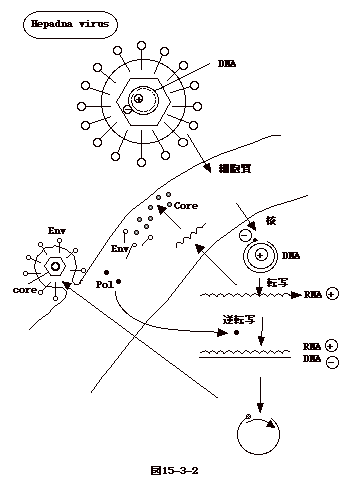
15-3-2:ヘパドナウイルス
Hepadnavirusの仲間で重要なのは、B型肝炎ウイルス(HBV)である。成人への感染では、持続感染に移行することは少ないが、母子感染では9割位が持続感染に移行し、慢性感染、肝硬変、或いは肝がんとなる。主な感染経路は血液(輸血)で、その他性行為でも感染が起こる。輸血によるHBV感染は、感染を調べるスクリーニング法が開発され、全ての輸血用血液を検査するようになってから殆どゼロになった。しかし、針刺し事故などで医療従事者にも感染したケースは時折見られる。酵母で作成した組替えワクチンが感染予防に有効であり、母子感染防止、医療事故感染防止に使用されている。
15-3-3:レトロウイルスの複製に関する補足
1.レトロウイルスには宿主域があり、ウイルス上の特定の遺伝子と宿主の特定の遺伝子で決まる(Odaka
& Yamamoto, Jpn J Exp Med, 36, 23, 1966)。例えば、マウスにしか感染しないマウス白血病ウイルス(ecotropic
murine leukemia virus、 MLV)の場合、ウイルスのgag遺伝子の中のp30がウイルス側の遺伝子であり、マウスの第5染色体上にあるFv-1
locusがマウス側の遺伝子である。
ウイルスが増殖出来ない組み合わせでは、逆転写迄は感受性細胞同様進行するが、プロウイルスが細胞質から核に移行し組み込まれる間で、Fv-1の抑制がかかる。in
vitroの組み込み系でFv-1の影響が見られないことから、ゲノムの細胞質から核への移行に宿主のFv-1遺伝子産物が関与しているのではないかと考えられている。又、ウイルスのp30が
Fv -1 による宿主域を決めている事から p30蛋白はプロウイルスの核移行に関与していると推論出来る。即わち、ゲノムRNAと粒子中で一緒にいた
p30は、逆転写後もプロウイルスDNAと結合し核に移行しているという事になる。そうすると、実際核に移行するのは
Gag 蛋白の中の p30か Gag-Pol 蛋白中の p30かという問題が出て来る。プロウイルスの細胞質から核への移行の機構はレトロウイルスの未解決な問題の一つである。
2. retrovirusの感染は細胞増殖に依存する。HIVは静止期の細胞にも感染が成立するとされているが、増殖している細胞の方が感染効率は遥かによい。MLVの感染は完全に細胞増殖に依存する。例えば、接触阻害(contact
inhibition)により増殖の止まった単層細胞を一部ピペットの先で傷つけMLVを感染させると、細胞の剥がれた領域に回りだけ感染する(Yoshikura,
J. Gen. Virol, 6, 183, 1970)。では、必要とされる細胞周期はどこかと云うことになる。
感染した細胞を非感染細胞の上に播くと、そこから感染が広がる。これを感染中心(infectious
center)と云う。MLVを細胞に感染させ時間を追って感染中心の紫外線感受性を調べると、感染が成立しウイルスを放出し始めると紫外線感受性が急に下がる。つまり感染成立は感染中心が紫外線抵抗性となる事で判定出来る。何故感染が成立すると紫外線抵抗性になる理由は恐らく感染成立前の紫外線の標的は細胞の染色体DNAであり、感染成立後は転写されたウイルスRNAが標的分子となる為ではないかと考えられる。実際、retrovirusは紫外線抵抗性で、且つ、感染成立後の感染中心はRNAの紫外線感受性を高める5-fluorouracilを細胞に取り込ませる事により紫外線感受性が高くなる(Yoshikura,
Virology 48, 193, 1972)。
一方細胞周期と感染の関係を見るには全部の細胞が同じ細胞周期にあるような同調培養を使う必要がある。その為には、接触阻害によりG1期に止め、新鮮血清添加により増殖を誘導するとよい。このような系では、血清添加後10時間位でDNA合成が開始し20時間でDNA合成が最盛期を迎え、細胞分裂の最盛期は30時間頃となる。
このような培養系で、血清添加後、色々な時間でウイルスを感染させ、感染中心の紫外線感受性を時間を追って調べてみる。すると、紫外線抵抗性になる迄の時間はG1期を進みS期の最盛期に近付くに従って次第に短くなる。又、感染細胞を細胞分裂を抑える薬剤で処理すると処理した時間分紫外線耐性になる時間が遅れる。このような観察から、必要とされる細胞周期は細胞分裂ではないかと推測された(Yoshikura,
J. Gen Virol, 8, 113, 1970)。その後分子生物学的技法を用い追認されているが、その正確な機構は未解明のままである。
しかし、もし、細胞分裂がretrovirusの感染に必須であるとするとどのような機構が考えられるであろうか。
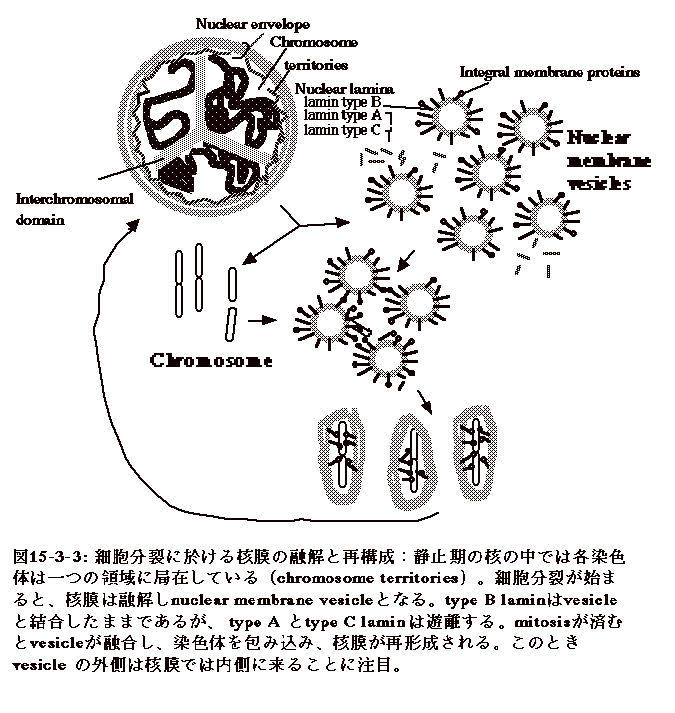
細胞分裂とはどういうものか、ここで少し考えてみる。細胞分裂は、核膜の融解が始まるprophase、染色体が細胞の中心に整列するmetaphase、染色体が次第に両極に引っ張られて行くanaphase、次いで分かれた染色体を取り込んだ核の出来るtelophaseと進行する。
prophaseで、核膜は融解しnuclear membrane vesicleとなり、同時に染色体、nuclear
lamina、等も遊離した状態となる。nuclear lamina由来のtype B laminはvesicleの外側に結合したままであるが、type
Aとtype C laminは遊離する。染色体が両極に分かれanaphaseに入ると、nuclear
membrane vesicleはlamin等と共に染色体を取り込みつつ融合し核膜を再構成する(図15-3-3)。