�@�t�@�[�W�̌�����U��Ԃ�ƁA���̌��ɂ����Ă��݂��ɑ��e��Ȃ��A�Q�̍l���������������ɋC�t���B�@�t�@�[�W���ŏ��Ɏ����I�Ɉ������̂̓J�i�_�n�t�����X�l��
d'Herelle�i1917�j�ł���B�ނ́A�t�@�[�W�͍ۂɊO����iexogenous�j��������E�C���X�ł���ƔF�����Ă����B����A�����������ւ��Ă����x���M�[��
Bordet
�́A�����̍ۂ͑��̋ۂɊ�������t�@�[�W�Y�o����\�͂���݁iendogenous�j����ƍl���Ă����B
�@����́A�O�҂� T4 �� T7 �Ȃǂ� virulent phage
�������A��҂͂��̏͂ŏq�ׂ� lysogenic phage�������Ώۂɂ��Ă����ׂł���B���݂��ɕʂ̃t�@�[�W���������Ă������ɋC�t�����Ad'Herelle
�́A�uBordet
�̓t�@�[�W���q�̍��������ۂ��������Ă��邩�炠��Ȍ��_���o���̂��v�ƍl���Ă����B
�@�t�@�[�W�̌������番�q�����w�����グ��Delbrück
�̃t�@�[�W�O���[�v�͑O�҂̃t�@�[�W��ΏۂɑI�ь������\�z���čs�����ׁAlysogeny�i�n�����j�̌��ۂ͂Ȃ��Ȃ����̃O���[�v�ɔF�߂��Ȃ������B�iJ.
Cairns, G.S. Stent, S.D. Watson Phage and Origins of Molecular Biology, 1996, CSHLQB�j�B�n�����̌��ۂ̖{�������߂Ė����ɂ����̂̓��V�A����t�����X�Ɉږ�����
Lwoff�i1950�j�ł���B
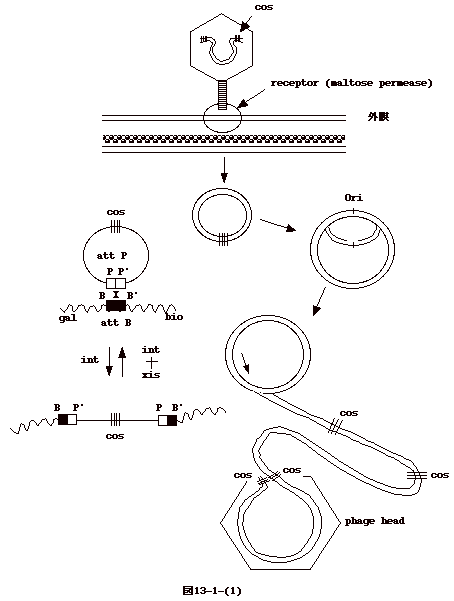
13�|1�FDNA ����
�@�����_�t�@�[�W�̓E�C�X���X���q�̒��ł͒����Q�d�� DNA
�̌`���Ƃ��Ă���B���[�� cohesive end�Ƃ������ʂ�����A���݂�����I��12
������P�{���ƂȂ��ĂQ�d�������яo���Ă���B��яo���Ă��鍽�͂T'���ł���B
�@�咰�ۂɊ�������ƁAcohesive end
�͂��݂��ɐ��f�������A�����_ DNA �͊�ɂȂ�BOri
���獶�E��������DNA �������i�s����i��type replication�j�B
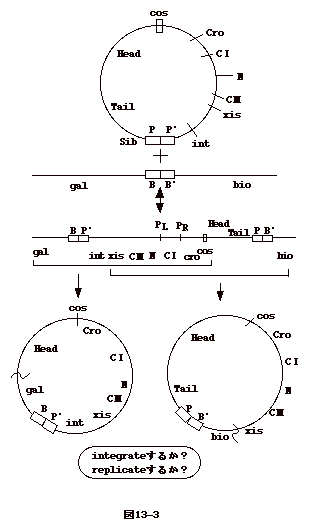
�@�E�C���X���q���o����n�ۃT�C�N���ł́A�₪�āArolling circle
replication ���n�܂�A�����_ DNA���������ɑ�R�Ȃ����� concatamer
���o����B�t�@�[�W�`���� cos ��F�����A�����_ DNA
���P���j�b�g�Ât�@�[�W�̓��ɋl�ߍ���ōs���B
�@����A�n�ۃT�C�N�C���ɓ��炸�A�����_ DNA
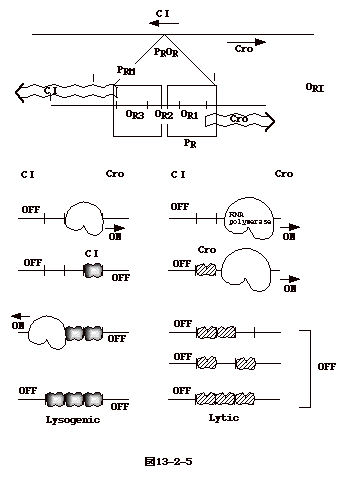
���咰�ې��F�̂ɑg�ݍ��܂��T�C�N�������ꍇ������B�����n���T�C�N���ilysogenic
cycle�j�Ɖ]���B���_ DNA �� att
�Ɖ]���ꏊ�𗘗p�����F�̂ɑg�ݍ��܂�A���̌�咰�� DNA�̈ꕔ�Ƃ��ĕ��������B�咰�ې��F�̏�̑g�ݍ��ݕ��ʂ�
gal �� bio �̊Ԃ�attB site �ɂقڌ��肳��Ă���B �t�@�[�W���iattP�j���咰�ې��F�̑��iattB�j���g�ݑւ����ʂ����܂��Ă���̂ŕ��ʓ��ّg�݊����̈�Ɖ]����BattP��
attB �ɂ� AT �̑������ʂ� 15 ���������i�}13-1-(1)�j�B
�@�g�ݍ��݂ɂ̓����_�t�@�[�W�� int
�`���i�{�h��`���j���K�v�Ƃ����Batt
�̓����_��`�q�n�}�̂قڐ^���ɂ���B�]���ăE�C���X���q���ł͍��̔��Α���
cos
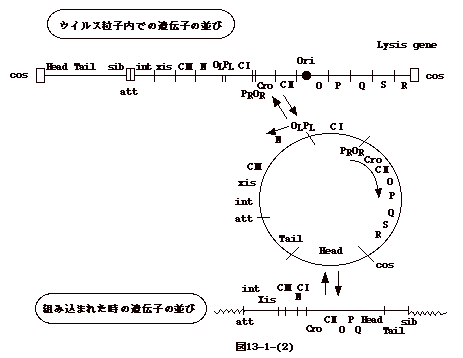
�ߖT�ɂ�������`�q�i�t�@�[�W����`�q�Ɨn�ۈ�`�q�q�j�͑g�ݍ��܂ꂽ�����_��`�q�ł͗��킹�ɂȂ�A�E�C���X���q���̃Q�m����att
�����ݗ����Ă�����`�q�iint �� sib�j�͗��[�ɗ����B���̗l�ȑg�ݍ��݂̃��f�����o�����̂�
Campbell �ł���i�}13-1-(2)�j�B
�@���O���ȂǑ咰�ۂ̐������������悤�Ȏh���ɂ��A�t�@�[�W
DNA
�͐�o����A�n�ۃT�C�N���ɓ���B���������A���v�D����l�Y�~�������o���悤�Ȃ��̂ł���B��o���ɂ́A�t�@�[�W��int
�� xis �`���i�{�h��`���j���K�v�Ƃ����i�}13-1-(1)�j�B