第10章 プラスミドと薬剤耐性
|
|
|
|
| 第9章でプラスミドについて簡単にふれたが、ここでは、複製と伝達について要点を記す。 |
|
10−1:複製調節
プラスミドは大きく2群に分けられる。
(1)ゲノムサイズが概して小さく(10〜30kb)細菌あたり10〜40コピーあるもので、ColE1、多剤耐性 因子R6Kなど、
(2)ゲノムサイズが概して大きく(60〜100kb)細菌あたり1から3のコピーしかないもので、F
因子や、多剤耐性因子 RP4、 R6、 R1 など。
プラスミドは細菌が二分裂するとき、両方の娘細胞に分配されなければ菌の中で存続出来ない。(1)と(2)のカテゴリーでその機構が違う。
(1)は、ともかく沢山のコピーが菌の中にあれば、確率的に1コピー位は必ず両方の細胞に行くであろう、という良い加減な(relaxed)機構を利用している。クロランフェニコル(CM)の存在下でも複製が起こり、菌を
CM
処理することで菌体あたりのプラスミド収量を増加出来る。遺伝子工学で非常に良く使用される
pBR322 もこのタイプである。
(2)は、細菌の染色体同様、プラスミドが厳密(stringent)に2つの細胞に配分される機構があると考えられるものである。CM
存在下で複製が起こらない。
プラスミドの伝達にはその為の遺伝子が必要なので、その遺伝子の無い
ColE1
などは、自分だけでは伝達され得ない。しかし、伝達遺伝子を持つプラスミドに助けられ伝達されることはある。他方、自己伝達はおろか他のプラスミドに助けられても伝達出来ないプラスミドもある。 |
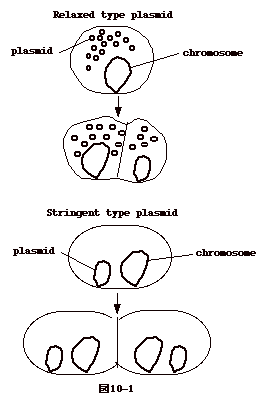
|
10−2:Relaxed Control
を受けるプラスミドの複製調節
|
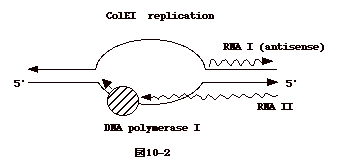
| ColE1は宿主由来の安定性の高いポリメラーゼで複製される。DNA
合成は常に RNAのプライマーを必要とするが、この場合も例外でなく、合成された
RNAII をプライマーとして DNA 合成が始まる。DNA合成開始は RNAI
により負の調節を受ける。RNA I はRNA IIとは逆向きに合成されるアンチセンスの
RNAで、これが RNA IIにhybridizeすることにより、その構造を変化させDNA合成のプライマーとしての機能を抑える。DNA合成は大腸菌DNAのような両方向ではなく一方向性である。 <問い>DNA
複製開始点構造が似た2種のプラスミドは1つの菌の中で共存し複製されない。これを、incompatibility
という。RNA I が関与すると云われている。その機構を考えよ。 |
|
 |
10−3:Stringentな複製制御を受けるプラスミドの複製調節
|
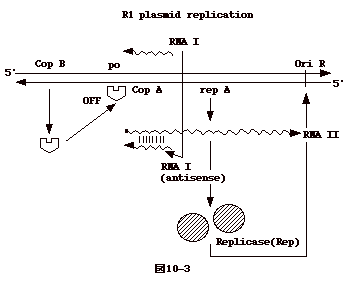
| R1 プラスミド
では、プラスミドのコードする不安定な RepA
蛋白により複製開始が調節されている。複製開始点は OriRである(接合伝達の
DNA 合成は、別の場所にある OriT から開始されるので注意)。RepA
蛋白はその遺伝子の乗っているプラスミドの OriR からのDNA
合成にのみ関与する(cis control )。 RepA 蛋白は RNA II
から翻訳される。RNA II はレプレッサーである copB
による負の調節を受けている。それと同時にRNA II の翻訳はRNA II
と逆の極性を持つ copA の産物であるRNA Iによっても負の調節を受けている。
以上、ColE1, R1何れの複製調節に於いても、逆の極性を持つRNA(anti-sense
RNA )による発現調節がある事に注意したい。 |
|
 |
10−4:伝達調節
薬剤耐性プラスミドを持った菌と持たない菌を混ぜると、耐性因子が伝達され感受性菌は耐性を獲得する。どうして、世の中の菌は全部耐性菌だけになってしまっていないのだろう。
プラスミドは菌が普通の条件で生育するには不必要である。そればかりでなく、それだけ余分なDNA複製のエネルギーが必要なので、生育には不利でもある。
プラスミドには、薬剤耐性や病原性を決める遺伝子が乗っている。いずれも、菌が自分に不利な条件で生き残る為に必要な性質である。そこで、そのような遺伝子は、菌の一部の集団に保存され、一旦、条件が悪くなればそれが全体の集団に伝達されるのが理想である。
このような目的に適合する現象が2つ知られている。
1つは plasmid elimination
である。プラスミドは、染色体と独立して複製されるので、脱落する事が出来る。特に、上の分類で、ゲノムの大きい(2)のカテゴリーのものは、エチヂウムブロマイド存在下で増殖させると、プラスミドを失う。
第2の機構は、やみくもにプラスミドが伝達されないようにする機構で
fertility inhibition と呼ばれる。
R100 プラスミドの接合伝達は一群の tra
遺伝子のコードする蛋白により行われる。この遺伝子群には、接合に必要な
pili の遺伝子も含まれている。 |
|
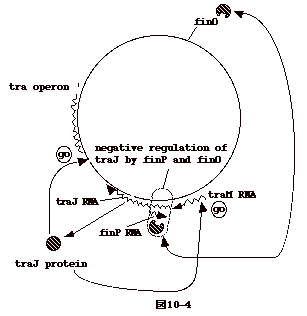
| tra 遺伝子は traJ
遺伝子産物により正に制御されており、traJが転写されるか否かで、接合伝達の可否が決る。 traJ
は、それとは逆向きに転写される finP
の遺伝子と5’側で重なっている。つまり、finP RNAは traJ mRNAのanti-sense
RNAである。finP RNAは全く離れた遺伝子座にある finO の産物とcomplexを作りtraJ
の翻訳を抑える。
R100 plasmid が伝達された時、受け取った側の菌には finO
産物はない。しかし、時間が経つと、R100 plasmidからコードされるfinO
産物が菌体に溜る。すると、finO産物によりtraJ
の翻訳が抑えられ、接合伝達が低下する。プラスミドを受け取った直後の菌が接合伝達能が高く、時間が経つと低くなるのはこの為である。 |
 |
<問題>F因子には finO
遺伝子が無い。ただし、finP 以下 tra 遺伝子は皆そろっている。
(1)オス(F+)の菌は fertility inhibition の現象を示すか?
(2)オスの菌に R100
を接合伝達すると、そのオス菌の接合能はどうなるか?
10-4bis: プラスミド維持機構
プラスミドが必要以上に伝達しない事も必要であるが、これが細胞分裂で、容易に失われるようでは、プラスミド自体存続出来なくなる。この機構として、toxin-antitoxin(TA)遺伝子がある。これは、細胞分裂の際に、プラスミドを受け取らなかった菌が選択的に死或いは障害に至る現象で、プラスミドの遺伝子が支配している。つまり、プラスミドが無いと菌の生残に不利になる機構をプラスミドが持っている、と云うことである。
TA系は菌を殺す毒素(T)遺伝子とその作用を相殺する抗毒素(A)遺伝子からなり、TとAとは遺伝子領域が重複している事が多い。その機構には2種類あり、一つはA遺伝子がanti-sense
RNAでT遺伝子の翻訳を抑える場合(Type I)であり、他は毒素が安定で、これを安定性の悪い抗毒素が結合し中和している場合である(Type II)。
Type Iの代表は上述のR1プラスミドのhok-sok遺伝子である。 hokは細胞膜を標的とする毒素をコードするが、そのmRNAは転写直後には翻訳されず3’側から消化され短くなって始めて翻訳可能となる(λファージのretro-regulationに似ている)。処が、sokのコードするmRNAは、hokのmRNAの5’領域と逆向き(anti-sense)塩基配列を持ち、短くなったhokのmRNAに結合する。出来た二重鎖RNAはRNase
IIIで壊され、プラスミドを持った菌は自分自身を殺す毒素をコードする遺伝子をもっているのに、生き残る事になる。一方、プラスミドを受け取らなかった場合、hok
mRNAは短くなって始めてsok mRAと結合する事、つまり長いmRNAである限り安定である事、hok、sok
それぞれのRNAの崩壊速度の違い等が原因となって、転写可能なhok mRNAが残り毒素がどんどん作られて菌は死ぬことになる。
Type IIの例としては同じR1プラスミドのDnaBを標的とするKid毒素に対するKis抗毒素など多くの例が知られている。プラスミドを受け取らないと、それぞれのmRNAの供給もなく、後は、毒素と抗毒素の残存で菌の生死が決まることになる。毒素の方が抗毒素より安定なので、プラスミドの無い菌には毒素だけ残り死ぬ(F.
Hayes, Toxins-Antitoxins: Plasmid Maintenance, Programmed Cell Death, and
Cell Cycle Arrest, Science, 301, 1496-1499, 2003)。
TA遺伝子の系は、菌が自分自身を殺す遺伝子を持っていると云う点で後に述べるapoptosisに似ている。
|
|
10−5:Hfr と F'
|
|
|
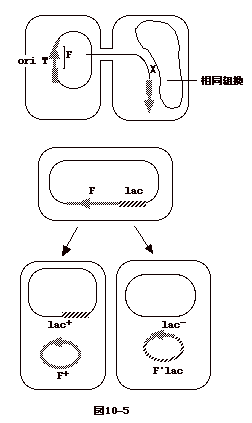
プラスミドは菌の染色体に組み込まれる事が出来る。プラスミドと菌の染色体双方にある
transposon の塩基配列を利用して組み込まれる。
F 因子が染色体に組み込まれると Hfr
となる。結果、染色体はあたかもプラスミドの一部であるかのように伝達される。 伝達の際の
DNA複製は OriT から1方向に起こる。従って、菌の DNA複製に要するほぼ2倍の時間で、伝達
DNA
複製が完了する。故に、大抵は伝達の途中で伝達が中断される事となる。OriT
は組み込まれた F 因子の中程にあるので、OriT
より前方のゲノムは伝達されないことが多い。つまり、メスはオスの染色体
DNA の一部は受け取っても残りのF 因子の DNA
までは受け取れないのでメスのままである。
受け取られたオスの DNA の一部はメスの DNAと相同組み換えを起こす。結果、オスの遺伝子マーカーがメスに伝達される事となる。
Hfr から F の正確な切りだしがおこれば、F+のオスになる。しかし、切り出しの時、非相同組換えにより、染色体
DNA の一部と一緒に F が出てくると、F' となる。F'lac は lac operon
がこのようなプロセスで F に組み込まれているものである。
<問>RecA−lac−のメスとF'lacをもつ菌を接合させた場合と、
RecA−lac−とHfr(lac+)を接合させた場合の違いを述べよ。 |
 |
10−6:Transformation
接合伝達は DNA が菌から菌へ pili
を介した接合により伝達される現象である。しかし、菌から菌へと遺伝情報が伝達されるには、DNAが菌に導入されればよい。
菌によっては、直接 DNA が菌に取り込まれ、その菌の DNA
と組み換えを起こし、その菌の性質を変える。これを、transformation
という。Avery の肺炎球菌の実験は、その例であり、DNA
が遺伝情報の担い手である事を証明した実験である。
DNA が感染性ファージ DNA やウイルスである場合transfection
と云う。
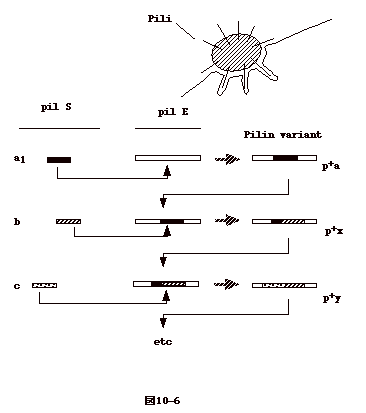
淋菌では pili の抗原性が変化し、それぞれの抗原性を持つ pili
遺伝子の一部が発現遺伝子座に持って来られる事が知られている。これは、一部の菌が溶菌し、そのDNAが他の菌に入るtransformation
による事が分かっている(図10-6)。 |
|
 |
|
|
|