8−1:遺伝子の存在の認識
我々は、或るものが存在するのかしないのかをどうやって認識しているのであろうか。我々の目に見えるものは、それだけで存在すると我々は理解する。
しかし、よく考えると、「存在」の認識は「非存在」が認識されて始めて、その認識にいたる。
日本人の目は黒い。これを決める遺伝子があるのかどうか。そのような疑問は、欧州人の青い目を見て始めて出てくる。SM
耐性遺伝子の存在はSM
感受性の菌と耐性菌があって始めてその認識に至る。
遺伝学は、この様な「変りもの」にヒトが興味を持ったことから始まる。エジプトのスフィンクスはそういう興味から生まれた幻想ではないだろうか。実際、そのような興味は中世から盛んに行われた同種異種の動物と動物のかけあわせ、さらには遺伝学へとつながっていく(F.
Jacob, Logique du vivant)。
遺伝学は「変りもの」の学とも云える。遺伝学は従って変異株の分離に始まる。「変異株分離」と単なる「混じりものからの選択」をまず区別しなければならない。
変異株の分離は遺伝的に均一な集団を得る処から始まる。画線培養によるコロニー分離はまさに遺伝的に均一な集団を得る為のものである。
遺伝学において、研究対象の遺伝的均一性は最も重要な前提である。
病巣から菌を分離し、何という菌による病気かを調べる菌の同定も一種の遺伝学である。2種の菌が混じっていては、同定は出来ない。Aの菌がlactoseを分解し(Lac+)maltose
非分解(Mal−)で、Bの菌が Lac−Mal+とする。もし、同定する菌がABの混じりであれば、結果は
Lac+Mal+のAともBともつかない別のCという菌として同定されるであろう。
8−2:進化
生物は生命が地上に生まれて以来進化を続け現在の多様な種の形成に至った、と考えられている。
生物が進化を続けるには遺伝子が常に変る事が必要である、進化は「遺伝子のデタラメな変異」と、その時の環境に適した「生物の選択淘汰」によるとされている。
進化とは、環境に生物が適応するように表現型を変え、これが遺伝し(云い換えると、表現型が逆に遺伝子を変え)、新しい種を形成する、と云うのが、いわゆるラマルキズムである。革命直後のロシアにおいて、ミチューリン、ルイセンコ等がこれに近い学説を唱え、スターリン政権に近付き、これを批判する
Vavilov などの遺伝学者を追放、死に追いやった。
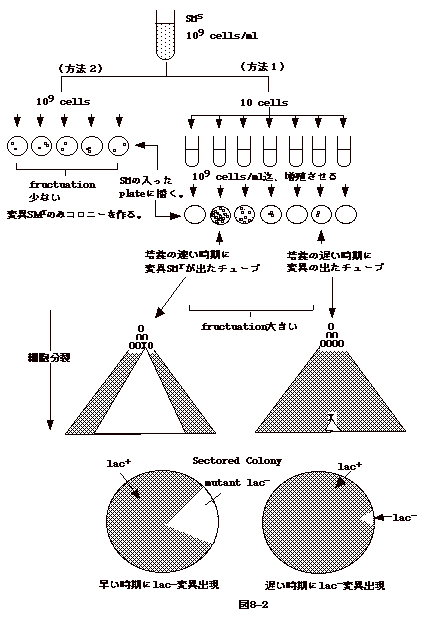
上の議論に関係して細菌学でも同様な議論があった。いま、ストレプトマイシン(SM)に感受性の細菌を
SM 存在下で飼育する。そうすると、SM耐性の菌が現れる。耐性菌は
SM により誘導された変異なのか?或は、SM
存在如何によらずそのような変異が起こり、それを選択したのか?
この疑問に対しなされた実験がDelbrückとLuria のfluctuation
試験である。
SM 感受性の菌を 109 匹/ml 用意する。これを10匹/ml
に希釈し 30 本の試験管に分注し、109 匹/ml迄増殖させる。これを、SM
の入った寒天平板に1 ml
づつ植えコロニーを作らせる(方法1)。片や、初めの109
匹 /ml の菌を1ml づつ 30 枚の SM入り寒天平板にうえる(方法2)(図8-2)。