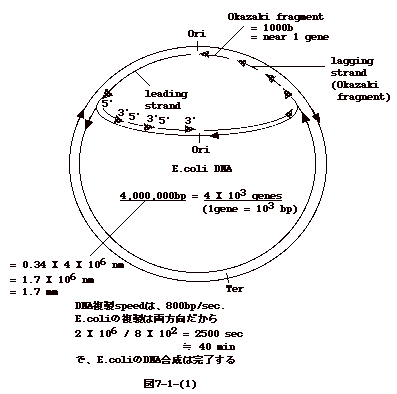
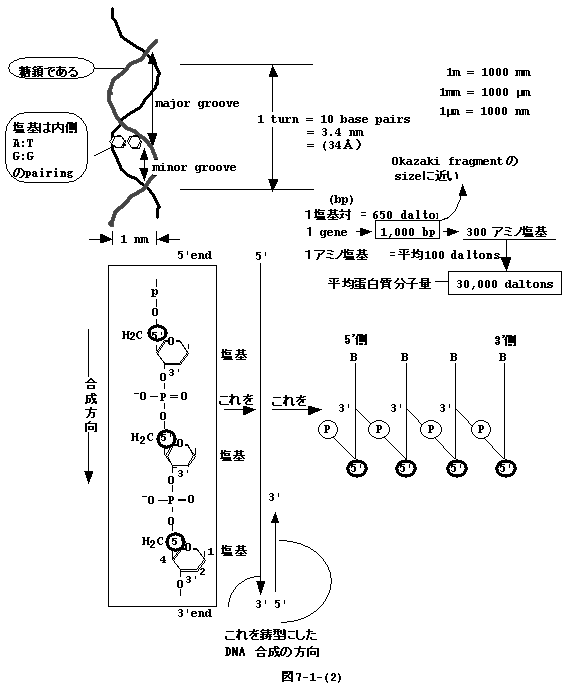
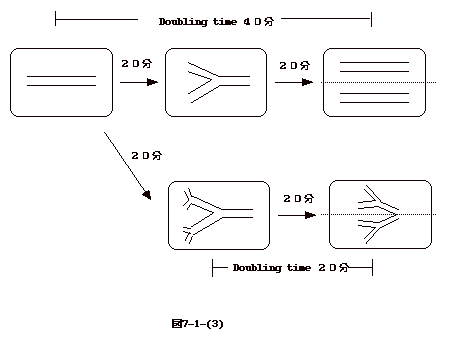
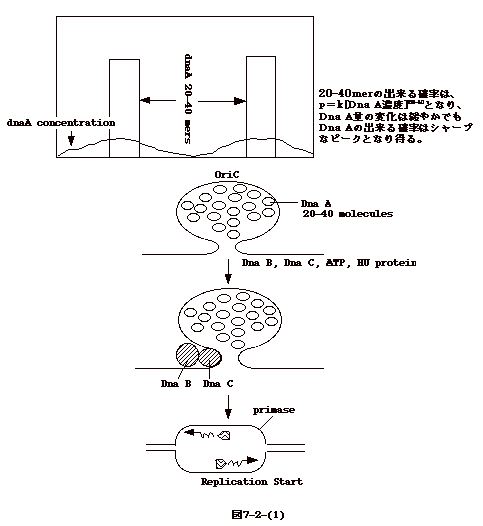
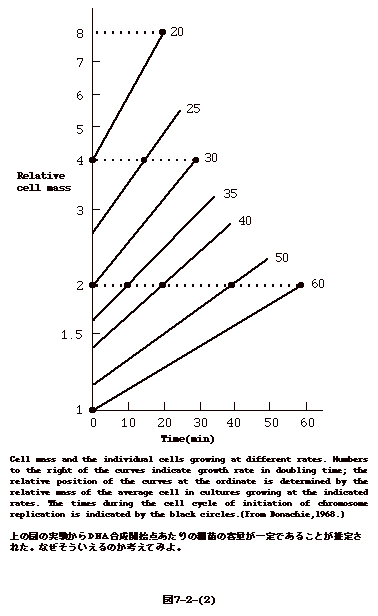
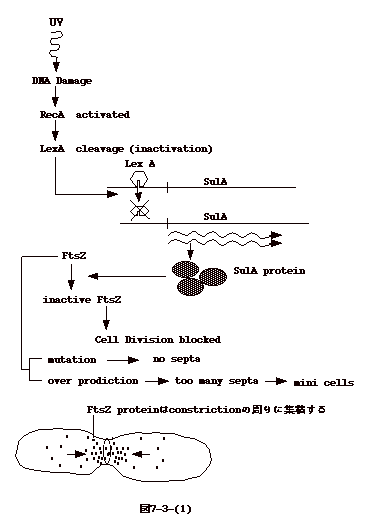
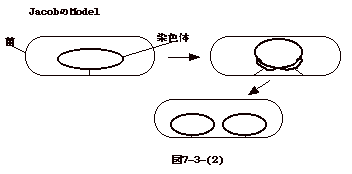
丂
忋偺幚尡偱丄拲堄偟偰栣偄偨偄偺偼丄DNA崌惉偑婲偙傞応強偼屌掕偝傟丄DNA偺曽偑庤孞傝婑偣傜傟側偑傜暋惢偟偰峴偔偲塢偆揰偱偁傞丅DNA崌惉偵娭傢傞抈敀暋崌懱偑嫄戝側暘巕偱偁傞偙偲傪峫偊傞偲丄榖偑椙偔暘偐傞丅揮幨偺応崌傕摨條偱丄RNA億儕儊儔乕僛偑DNA偺忋傪擲鈌偺嵶朎幙偺傪偐偒暘偗摦偄偰峴偔傛傝偼丄DNA傪庤孞傝婑偣偰mRNA傪崌惉偟偰偄偔曽偑崌棟揑偺傛偆偵巚傢傟傞丅
丂DNA偺暋惢揰偑妘暻偺弌棃傞応強偵屌掕偝傟偰偄傞偺偼傛偄偑丄偙傟偩偗偱偼暋惢偟偨2偮偺DNA僐僺乕偑俀偮偺柡嵶朎偵偦傟偧傟暘攝偝傟傞偙偲傪曐忈偱偒側偄丅恀妀嵶朎偺愼怓懱偼centromere偵晅偄偨kinetochor傪夘偟偰嵶朎偺椉嬌偵暿傟傞偑乮恾16-3-6-(5)乯丄嵶嬠偵傕帡偨婡峔偺偁傞偙偲偑暘偐偭偨丅
瀰嬠偺Bacillus subtilis偱RacA曄堎乮RecA偱偼側偄両乯偼朎巕宍惉偺堎忢傪棃偡偑丄偦偺尨場偼DNA偺攝暘堎忢偱偁傞丅RacA抈敀傪Chromatin
immunoprecipitation偲塢偆媄朄偱挷傋偰傒傞偲丄摿偵DNA暋惢拞怱偵嬤偄張偵偁傞centromere椞堟偲寢崌偟偨丅Centromere偵寢崌偡傞RacA偼丄瀰忬傪偟偨嬠偺椉嬌偵嬊嵼偡傞DivIVA偵寢崌偟偨丅懃偪丄愼怓懱偺DNA暋惢奐巒揰偵偁傞centromere偼RacA傪夘偟嬠偺椉嬌偵偁傞DivIVA偵寢崌偡傞偙偲偵側傞丅偦偆側傞偲丄2偮偵暘楐偟偨愼怓懱偼偦傟偧傟侾働強偱嬠偺椉嬌偵晅偄偰偄傞偺偱丄嵶朎暘楐偑婲偙傟偽丄愼怓懱傕俀偮偵暿傟傞偙偲偵側傞丅
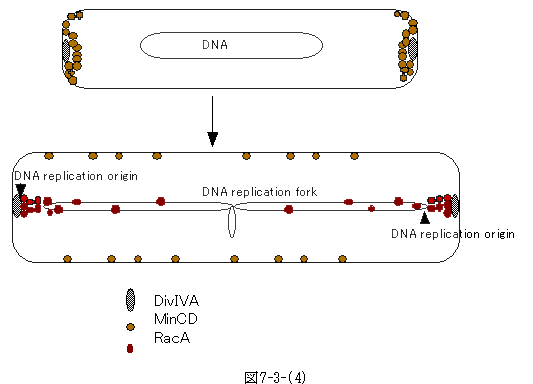
丂偟偐偟丄偙傟偱偼丄愼怓懱偺暋惢奐巒揰偼嬠偺嬌偵晅拝偟偨傑傑偵側傝丄DNA暋惢偵嵎偟巟偊傞丅偙傟傪惂屼偟偰偄傞偺偑MinCD偱丄嬠偑暘楐偵岦偗偰惉挿偟偰偄傞娫偼丄MinCD偑DivIVA偺嬌嵼偡傞晹暘偵廤傑傝丄嵶朎暘楐偺奐巒傪梷偊丄摨帪偵RacA偺DuvIVA傊偺寢崌傕梷偊傞偲偄偆巇慻傒偲側偭偰偄傞丅壖愢偺晹暘傕偁傞偑丄Jacob偺壖愢傊偺堦偮偺夞摎偱偁傞乮S.Ben-Yuda,
DZ. Rudner , R. Losick, RacA, a bacterial protein that anchors chromosomes
to the cell poles, Science, 2003, 299, 532-536乯丅
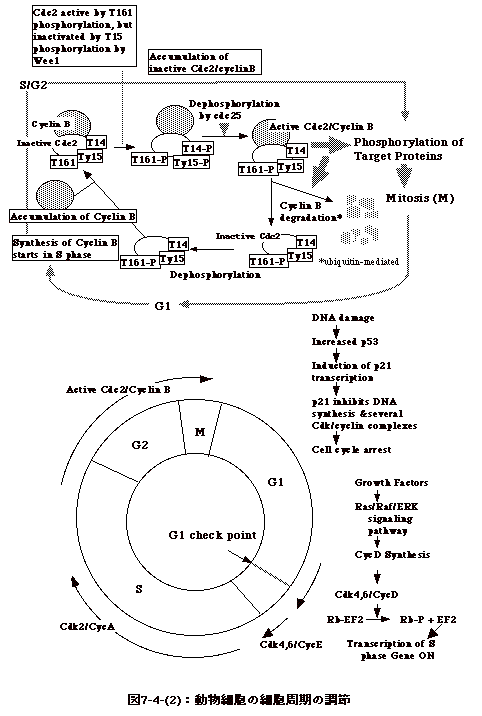
7亅4丗嵶朎廃婜
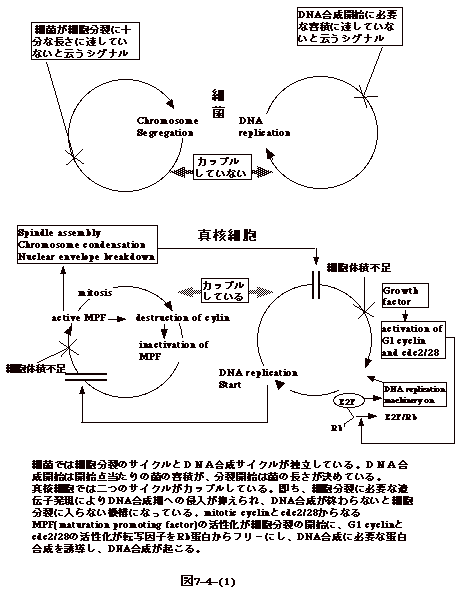
丂嵶嬠偁傞偄偼摦暔嵶朎偺憹怋偼丄DNA偺暋惢偲嵶朎暘楐偺俀偮偺尰徾偐傜側偭偰偄傞丅
丂恀妀嵶朎偱偼嵶朎暘楐偐傜嵶朎暘楐傑偱偺帠徾丄嵶朎廃婜丄偼丄G1丄S乮DNA崌惉婜乯丄G2丄M乮嵶朎暘楐乯偑丄偙偺弴彉偱惓妋偵恑傓丅M婜偑廔傢傜偢偵師偺S婜偵擖傞帠偼側偄丅
丂嵶嬠偱偼丄嵶朎暘楐偑嵪傑側偄偺偵師偺DNA崌惉偑巒傑傞丅偮傑傝丄恀妀嵶朎偱偼乽嵶朎暘楐傪偟偰偄傞偲塢偆乿僔僌僫儖偑DNA崌惉婜傊偺僐儈僢僩傪梷偊丄DNA崌惉傪偟偰偄傞偲塢偆僔僌僫儖偑嵶朎暘楐傪梷偊偰偄傞偑丄嵶嬠偱偼丄偦偺傛偆側僔僌僫儖偵傛傞嵶朎撪偺枾愙側挷愡偼側偔丄憹怋偺懍偄嬠偱偼丄嵶朎暘楐偲DNA崌惉偑摨帪偵婲偙傞乮恾7-4-(1)乯丅偨偩偟丄婛偵弎傋偨傛偆偵丄DNA崌惉奐巒偐傜堦掕帪娫屻偵嵶朎暘楐偑婲偙傞偺偱姰慡偵DNA崌惉偲嵶朎暘楐偑彑庤偵婲偙傞偲塢偆偙偲偱偼側偄丅
丂偙偺傛偆側忬嫷傪斀塮偟丄恀妀嵶朎偱偼悢懡偔偺堚揱巕偑嵶朎廃婜偺挷愡偵娭梌偟偰偄傞丅
丂恾7-4-(2)偼摦暔嵶朎偺嵶朎廃婜傪揨傔偨柾幃恾偱偁傞丅嵶朎廃婜偺挷愡偵偼俀偮偺僠僃僢僋億僀儞僩偑偁傞丅DNA崌惉偵擖傞偐偳偆偐乮偮傑傝G1偐傜S婜傊偺堏峴乯偲嵶朎暘楐傪巒傔傞偐偳偆偐乮G2偐傜M婜傊偺堏峴乯偺俀揰偱偁傞丅