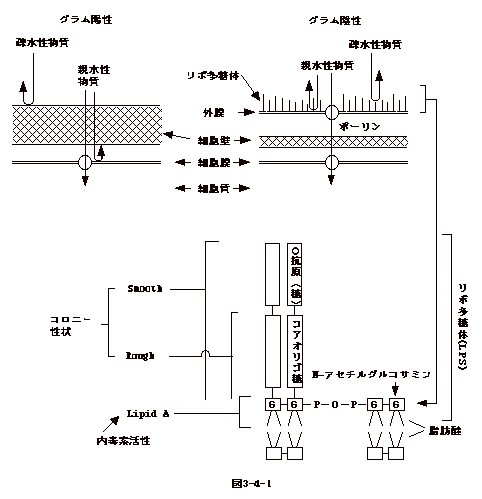
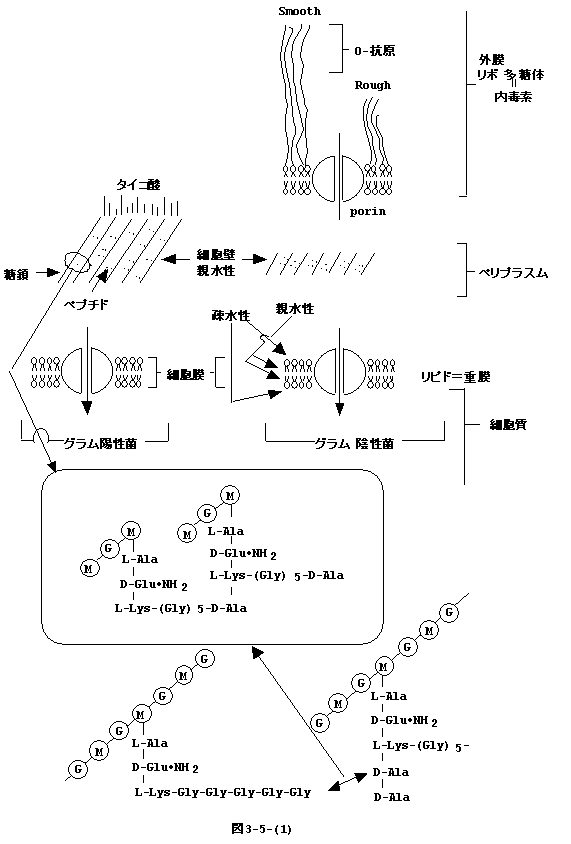
GlcNAC−MurNAC−pentapeptide−(Gly)5は
carrier lipid
から細胞壁のペプチドグリカンに受け渡される。しかし、これではGlcNac-MurNac-pentapeptide-(Gly)5が繰り返す糖鎖が出来るだけである(図3-5-(1)左下)。 ペプチドグリカンが丈夫な編目を作るにはペンタペプチド間の架橋が必要である。架橋は、グラム陽性菌では(Gly)5、グラム陰性菌では
diaminopimelic acid が別の鎖のD-Ala−D-Ala
と反応し完成される(図3-5-(1)右下)。もし別の鎖のD-Ala−D-Alaでなく同じ鎖のD-Ala−D-AlaであるとGlcNAC-MurNACの繰り返しの糖鎖はお互いに連結されず、丈夫な細胞壁は出来ない。どの様な機構が同じ鎖と別の鎖を識別しているのだろうか。
唾液などに存在するlysozymeは糖鎖を切る。ペニシリンなどの抗生物質は上のプロセスの何処かを抑える。Cycloserine
は L-Ala から D-Ala への変換と、D-Ala のトランスペプチデーションを抑える。Vancomycin はGlcNAC-MurNAC-pentapeptide-(Gly)5のcarrier
lipid から細胞壁への受け渡しを抑える。
ペニシリン(penicillin)、セファロスポリン(cephalosporin)は架橋を抑える。ペニシリンは構造的に
D-Ala−D-Ala
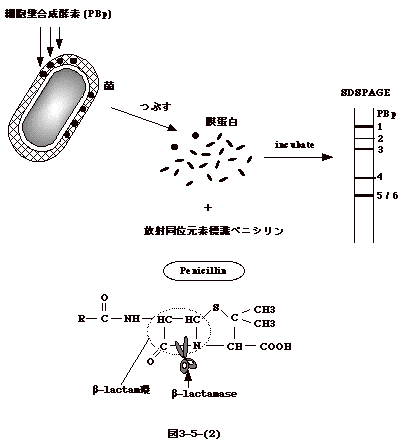
に似る。つまり、架橋酵素の基質に似ている。従って、アイソトープ標識したペニシリンと細胞膜分画を混ぜ、放置してから電気泳動すると、架橋酵素が標識されたバンドとして見えてくる(図3-5-(2))。このように見えてくる蛋白をペニシリン結合性蛋白(PBP)と云う。メチシリン耐性となったブドウ球菌(MRSA)では感受性菌にない新しいバンド(PBP2')が出現する。
狭域 penicillin
はグラム陽性菌、一部のグラム陰性球菌、梅毒にしか効かない。グラム陰性菌には外膜があるため多くの場合penicillin
がこれを通過せず、細胞壁合成部位に届かない。
広域 penicillin
は外膜を通過し、グラム陰性菌にも有効である。ただし、どの広域
penicillinもブドウ球菌の penicillinase(ベータラクタマーゼ)で壊される(図3-5-(2))。
cephalosporin は penicillin
同様ベータラクタム環を持つ。ブドウ球菌のベータラクタマーゼでは壊されないが、多くのグラム陰性菌のベータラクタマーゼで壊される。半合成により,広い抗菌スペクトルを持った
cephalosporin
が開発され、第一(セファゾリン、セファレキシン)、第二(セファマンドル、セフォテタン)、第三世代(セフォタキシム、セフォペラゾン)と呼ばれる。第三世代のほうが抗菌域は広いがグラム陽性球菌への抗菌力は落ちる。MRSA
(メチシリン耐性黄色ブドウ球菌)による病院感染で犠牲者が多く出たのは、第三世代の
cephalosporin
を全ての菌に効く万能抗生物質と医者が誤解したのが原因
である。ペニシリンの仲間にはこの他 carbapenem 系、monobactam
系のものがある。
<問い>グラム陽性菌をペニシリン存在下で増殖させると、細胞壁の無い菌になる。これを、protoplast
と云う。マイコプラスマはこのような状態に良く似ている(ただし、マイコプラズマの細胞膜はステロールを含むという点で他の細菌とは違う)。陰性菌では同様の処理をするとどういう構造になるか?グラム陰性菌で出来る構造はprotoplastとは異なり、これをspheroplast
という。
protoplast あるいはspheroplast を作るもう一つの方法は?
<問い>蛋白合成を止め、細胞分裂も止めるような抗生物質、例えばクロランフェニコル、存在下でグラム陽性菌をペニシリン処理したら、protoplast
は出来るか?