第2章 微生物 |
|
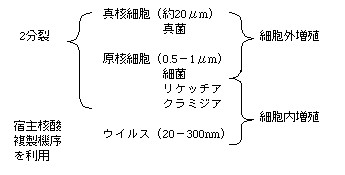
| 生き物は原核生物(prokaryotes)と真核生物(eukaryotes)に分けられる。前者には核がない。後者には核がある。 それぞれを宿主とするウイルスがいる。ウイルスは宿主外ではDNAかRNAいずれかが、蛋白質の殻に囲まれた形態をとる。ウイルスは宿主細胞に感染すると、宿主の遺伝子発現系をそっくり 利用し自分の遺伝子コピーとその蛋白を大量 に作る、従って、細胞に1コピーしかない遺伝子を研究するよりもウイルスの遺伝子のほうが遥かに研究が容易である。この為、ウイルス学は多くの細胞生物学の基本概念を生み出した。 次のようにまとめる。
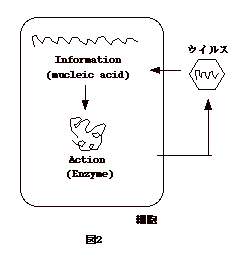
生物は増殖を続ける為に「情報を蓄える」機能と、「その情報に基づき働く」という機能を持つ。だからもし両方を揃えている細胞に情報さえ入れば、その情報は細胞の内でプロセスされ機能を発揮する。ウイルスは自分自身では「働く」という機能を持たない。 しかし、情報は持っているのでそれを細胞にインプットすれば自分を増やす事が出来る(図2)。
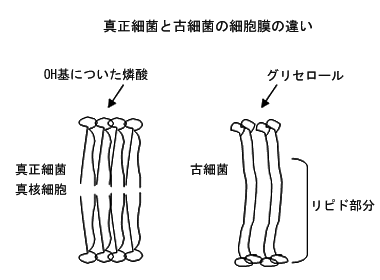
原核細胞のEubacteria(真正細菌)とArchaea (古細菌)の内、医学領域で問題になるのは真正細菌の方である。古細菌は沸点近い温度の温泉や岩塩等生物が住めるとは思えないような場所に生息する菌として1970年代の後半に報告された。ヒトに病気を起こすものは知られていないが、反芻動物の腸管でメタン発酵している古細菌に属するものがある。高温や高塩濃度など厳しい条件に限らず海水や土壌など環境中に広く存在することが知られるようになった。古細菌については、JL Howland, The Surprising Archea. Oxfordを読むと良い。古細菌の中には、四角や三角の結晶のような形態のもの、不定形のもの、泡がプクプク出ているような形態のもの等がある。又、古細菌は、高温を好むもの、メタン産生するもの、好塩性の3群に分けられる。 古細菌も真正細菌も、多くは球状であったり桿状であったりそう違わない。どうしてこのような形態のものが生物として基本的に違う群に入ることが分かったのであろうか。それは、遺伝子の解析による。しかし、古細菌と真正細菌の間には遺伝子交換もあり普通に遺伝子を比べただけでは結果は出る筈もなかった。 1977年、C. Woeseは、機能的に変化し難く遺伝子交換の影響も受けないリボソームRNAの塩基配列を解析することにより古細菌の一つであるメタン産生菌は真正細菌、真核細胞何れとも同じくらい進化的にかけ離れている事を示した。 進化の過程で先ず真正細菌と古細菌が分かれ、次いで古細菌と真核細胞が分かれた、と云われている。即ち、古細菌、真正細菌、真核細胞生物は遺伝子の相互の類似性に基づき、進化系統樹を書く事が可能である、しかし、ウイルスになると、ウイルス全体の遺伝的系統関係は見られなくなる。即ち、ウイルスはそれぞれの宿主の中で、独立してウイルスとして出現し進化した、と考えられる。これをpolyphylogeneticと云う。 真正細菌も真核細胞も細胞膜はフォスフォリピドの2重膜からなるが古細菌はリピドの1重膜で、細胞の中と外になる両端が親水性のグリセロールとなっている。考えて見ると、古細菌の生息する高温では2重膜では膜がお互いに剥がれて構造を維持出来ないかも知れない。
真核細胞生物は、原虫のような単細胞生物と回虫や人間のような多細胞動物に分かれる。多細胞生物の特徴は、分化した細胞が集まって社会を形成し、個体形成しているところである。従って、それぞれの細胞は、「個体の中で自分が何処にいるか」を認識し、それに応じ機能を発揮(所謂「分化」)しなければならない。即ち、この為には、個体の中での分化の場、或いは勾配(例えば頭から尻尾にかけての分化に関与する物質の濃度勾配)、の形成と、細胞と細胞との情報交換が必要となる。
このような単細胞が集合し多細胞形態をとる現象が、単細胞生物から多細胞生物への進化の過程を示すものとすれば、単細胞から多細胞への進化は、原核細胞、真核細胞がそれぞれ進化した後、独立に起こり得た現象で、真核細胞である事が多細胞への進化の要因ではないと考えられる。 多細胞化は上の様に単細胞が集合して出来ても良いし、分裂した細胞が一つの空間に閉じ込められバラバラにならない事でも可能である。水の中で多細胞化した微生物は後者に当たり(例えばボルボックスVolvox)、地上の微生物の場合は前者に当たる(例えば粘菌)、とBonnerは云っている(J. Bonner, First Signals, Princeton University Press, 2001)。 このような同じ細菌同士の相互作用だけでなく、口腔細菌に見られるような異なった細菌同士の共生、細菌と真核細胞との共生、など、学問領域として未開拓の分野が多くある。環境の中でも細菌、カビ、原虫、ミミズなど、思いがけない程多様な生物が相互に作用し生態系を作っている。微生物生態学は、微生物の環境浄化への応用などを考えると今後非常に重要な部門となってくる。又、微生物が病気を起こすのは、微生物と宿主の相互作用の結果である。種を越えた生物同士の相互作用の機構の解明は、医学領域で大きな研究分野、微生物生物学(Microbial cell biology)、として発展している。 |
|
 原核細胞にはEubacteria(真正細菌)とArchaebacteria
(古細菌)が存在するが、この二者は遺伝学的に隔たりがあり、生物をEubacteria
、Archaebacteria、Eukaryoteの3つに分ける考えが受け入れられている。
原核細胞にはEubacteria(真正細菌)とArchaebacteria
(古細菌)が存在するが、この二者は遺伝学的に隔たりがあり、生物をEubacteria
、Archaebacteria、Eukaryoteの3つに分ける考えが受け入れられている。