微生物遺伝学演習 |
|||||||||||||||||||||||||||||||||||||||||||
| 分子遺伝学の演習問題の幾つかは、米国タフツ大学医学部の分子生物学シラバスに載っている問題を基にしているが、難しすぎるので易しく改変した。 | |||||||||||||||||||||||||||||||||||||||||||
演習1.次の文章を読み、下の質問に答えよ。 |
|||||||||||||||||||||||||||||||||||||||||||
〈質問〉
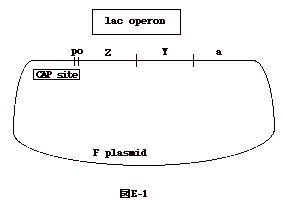
解説F'lacはplasmidにlactoseが組み込まれた。図E-1の様な構造をしている。repressor (R)をコードしているI遺伝子は一緒にいない。 |
|||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||
| しかし、F'lacを導入する大腸菌には、I遺伝子を持ったものもいるだろうと想像しておく。 Rは、lactoseがなければ lac operon の operator(0) にくっつき、この operon の発現をおさえる。lactose があるとその誘導体 allo-lactose が R と結合し、operator につかなくなり lac operon は ON になる。 lactose operon の発現は、lactose operon 上流の CAP site に、CAP(catabolite activator protein)が結合し、転写を促進することで、もう一つの調節を受けている。 CAP は glucose 存在下では CAP site に結合しない。これは glucose の細胞への取り込みにより、cAMP を作るアデニレートシクラーゼ活性がガタンと落ち、かつ、CAP は cAMP と結合して始めて CAP site に結合出来るからである。従って、glucose があると、正常の菌(wt)では lac operon はOFFになる。 問題1はwtと#1の lac operon の発現に何故 lactose が必要か、というのであるが、上の説明で明らかなように、wtと#1にはI遺伝子が正常に機能しているから、と推定出来る。 問題2は、wtと#2では何故 glucose が lac operon を OFF にするか、という問題である。これも上の解説通りである。 問題3は、#1が glucose の影響を受けない理由を聞いている。答えは一様ではないが、 glucose を取り込めないか、取り込んでもアデニレートシクラーゼが低下しないか、或いは、cAMPを壊す酵素(phosphdiesterase)が欠損しているか、要するに、glucose により cAMP の量が減らない変異株というのが答えである。 問題4は、何故#2は、lac operon の発現に lactose を必要としないか、との問いである。おそらくI遺伝子に欠損があるからである。 問題5は、#4がなぜ、如何なる条件下でも lac operon
の発現があまり高くならないか、と聞いている。F'lac
は正常な事を頭に入れておくと、これは cAMP に関係する事か、I遺伝子産物に関わる事と考えられる。 |
|||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||
| 問題6:グルコースの影響を受けるとwtと#2が
cAMP 添加によりグルコースによるlac operon の shut off
を防ぐ事が出来る。#1、#3いづれもグルコースの影響を受けない菌であるからcAMPの影響を受けない。
演習2.次の文章を読み、下の質問に答えよ。
(質問)
解説下の方の図で、始めの変異株の DNA の方が loop を作っている事に注目する。また、その長さが 1200bp である事からトランスポソンらしいと見当をつける。トランスポソンは Jumping gene と云われるように飛んで歩く遺伝子である。これが、Z 遺伝子の真ん中に飛んできたと推定する(問題1の解答)。そうすると、トランスポソンの中に転写停止シグナルがあれば、転写はそこで止まるので、Y,A を欠く mRNA が出来る(問題2の解答)。Type1 復帰変異は転写停止シグナル部位に変異が起きて、下流のY、Aまで読み取るようになったか、変異により新たな promoter が挿入 DNA に出来たかである(問題3の解答)。Type2 は正確なトランスポソンの切り出しである(問題4の解答)。Type3 は rho の変異でトランスポソン挿入によって出来た停止シグナルで転写がとまらなくなった、と考えればよい(問題5の解答)。
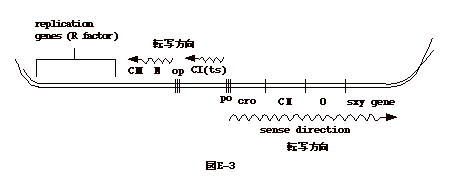
演習3.下の文章を読み、質問に答えよ。ワンダフル株式会社はベンチャー会社である。この会社は最近E-3図のようなベクター系を開発した。これを用い性転換蛋白(sxy)を大量に製造しようとしている。このプラスミドの一部は大腸菌R因子とラムダファージ由来であり、sxy遺伝子の転写調節領域は含まない。 |
|||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||
| ラムダファージの CI
は温度感受性なので、高温でレプレッサーは不活性化される。sxy
の合成は低温で停止し、高温にするとsxyの合成が再開する。
(質問)
解説問1:ラムダファージの promoter は cI 遺伝子を挟んだ PL とPR、cI をコードする mRNA の PRE、PRM、int をコードする mRNA の PINT があるが、ここでは場所から PR 以外無い。 問2:ラムダファージが菌に感染すると、N と cro を転写した処で転写が止まる。N とcro の蛋白だけが出来る訳であるが、N の遺伝子産物は転写停止を妨げ(anti-termination)、N と cro のより下流迄転写が継続するように働く。従って、sxy の発現には N 遺伝子を必要とする。 問3:低温で sxy 合成が止まり、高温で合成が起こるのは、cI が温度感受性の為である。cro,cII と読み取られると、cII 蛋白は、cI の promoter である PRE に働き、cI(レプレッサー)の転写を促進する。cI は PR から右側への転写を抑える。この為 cro 以下、下流の読み取りは OFF となる。 問4:これは、ラムダファージの遺伝子調節の上で最も理解しにくい部分である、cro と cI の微妙なバランスを理解しなければならない。PROR は cI および cro が結合する場所を3つ持っている。左から O3、O2、O1とする。cI は O1、O2、O3、の順に付いていく。つまり cI の量が増えれば一番左の O1 まで付く。cro は逆の順で付いていく。 上の問題では、高温なので cI は壊されており、このファクターは無視して良い。すると、O3、O2まで cro がついたのでは RNA ポリメラーゼは PR につき得るが、cro の量が増加しO1までつくようになると、ポリメラーゼがプロモーターにつけなくなる。すると、右側への写は OFF になる。従って、cro 遺伝子をノックアウトすると、sxy の生産は持続する。ラムダファージの感染サイクルを復習すること。
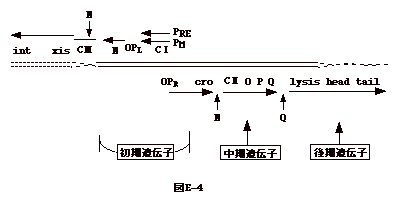
演習4.次の文章を読み、質問に答えよ。ラムダファージの感染しない大腸菌変異株がある。この大腸菌ではラムダファージの吸着、DNA の注入、初期遺伝子(N、cro)の転写迄は正常に起こるが、中、後期遺伝子の転写がない。この変異大腸菌は RNA ポリメラーゼに変異を持っている。この変異大腸菌でも増殖できるファージ変異株が分離された。この変異ファージは N 遺伝子に変異を持つ。しかし、N 遺伝子の変異ファージ総てがこの変異大腸菌で増殖できるわけではない。ラムダファージの遺伝子地図は図E-4のようになっている。 |
|||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||
(質問)
解説問1.N 遺伝子産物は anti-terminator なので正常な RNA ポリメラーゼとは反応し、左は cIII,xis,intと、右は cro,cII,O,P,Q...と転写を進行させる筈である。しかし、大腸菌の RNA ポリメラーゼが N 蛋白と反応出来ないような変異を持っていると、anti-termination は不可能である。 問2.変異RNAポリメラーゼと反応出来るようなN蛋白の変異が起きればよい。 問3.anti-termination 問4.N 及び cro の転写後の転写停止は rho 依存性である。したがって、rho を欠く菌では N 遺伝子に変異があっても、転写停止がおこらず、ファージは増殖出来る筈である。
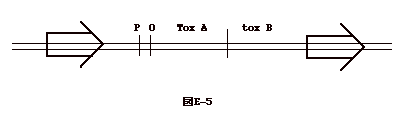
演習5.次の文章を読み質問に答えよ。大腸菌のある株はヒトや動物に下痢を起こす毒素を出す。この毒素は A と B のサブユニットから成り、それぞれ toxA、toxB の遺伝子がコードしている。toxA と toxB は一つの operon をなし、両側には同方向反復配列がある(図E-5参照)。 |
|||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||
菌は常に毒素を生産しているわけではないので、おそらく何かが inducer の役割を果たし制御が行われていると考えられる。 この遺伝子とは連鎖していない toxR 遺伝子に変異が起こると、毒素生産に影響が出てくる。toxR が欠損すると、toxA、toxB の転写が止まる。toxR 遺伝子の中の殆どの point mutation による変異は同様な効果を持つが、一部の point mutation では毒素があらゆる条件で生産され続ける。 毒素生産と同時に調節されている遺伝子は線毛(pilus)の遺伝子である。線毛は細菌が細胞に接着するのに重要な働きをする。上に述べた toxR 遺伝子変異は線毛遺伝子の発現に対しても toxA、toxB に対するのと全く同じ影響を与える。線毛遺伝子は toxR、toxA,toxB とは別の遺伝子座にある。 toxA、B 遺伝子の欠損は線毛の発現に影響せず、毒素生産も線毛発現に影響を与えない。 toxA、B の operon 内の変異には、毒素を常に発現し続けるようになるもの(constitutive)がある。この constitutive な発現は toxR に依存しない。また、この変異で線毛遺伝子発現が constitutive になる事もない。
(質問)
解説酵素には外毒素と内毒素がある事を思い出そう。外毒素は A、B の二つのサブユニットから成る事が多い。A(active)は活性を担い、B(bionding)は細胞への吸着を担う。 問1.明らかに、toxR は toxAB と pili の発現を促進する positive regulator である。おそらく、CAP のように RNA ポリメラーゼがプロモーターに結合するのを促進していると思われる。もう一つの可能性はプロモーターを選択するシグマ因子が考えられる。 問2.大部分を占める変異は toxR が不活性化されたものであろう。常に毒素を生産する変異は、蛋白の構造変化で常に toxAB オペロンを ON にするようになったものであろう。 問3.toxAB と pili の遺伝子は toxR を転写に必要とする似たプロモーターを持っている。 問4.toxAB のコード領域の上流に toxR に依存しないプロモーターが挿入されればよい(例えば、トランスポソン)。 問5.おそらく、繰り返し配列を利用して遺伝子コピーが増えたのであろう。
演習6.麻疹ウイルスは次の性質を持つ。
演習7.次の文章を読み、質問に答えよ。ある細菌学者が急に痴呆状態になり、これが M.ikarechattus の感染によるものと判明した。このM.ikarechattus に感染する二重鎖 DNA virulent phage が最近発見され話題となっている。 その phage は次のような性質を持つ。
(質問)
解説上の実験を纏めると、
問1.CM は蛋白合成阻害をする抗生物質である。従って、初期 RNA は宿主のすでに ある RNA ポリメラーゼ(蛋白)で転写されると考えると、説明がつく。 問2.Rifs の菌には Rif に感受性の RNA ポリメラーゼがあり、Rifr の菌には Rif 耐性 のポリメラーゼがある。従って、このファージは宿主の RNA ポリメラーゼを初期 RNA 合成に使用すると推定されている事から、初期 RNA も同様な感受性を示す。 問3.後期 RNA が CM 感受性なのは、初期に後期 RNA 合成の為の RNA ポリメラーゼ(蛋白)が合成されなければならないから、と推定される。 問4.問3への解答から後期 RNA 合成の為のポリメラーゼの一部はファージがコードしている、と考えられる。ファージ遺伝子69の変異の本体の可能性として、
問5.宿主のRNAポリメラーゼが温度感受性なので、もし、シグマ因子のみファージが 提供しているのであれば後期遺伝子は温度感受性であろう。(初期遺伝子は?)。もし、ファージがポリメラーゼそのものをコードしているのなら、後期遺伝子は宿主のRNAポリメラーゼの温度感受性にも関わらず温度に影響されないであろう。(初期遺伝子は?) 注:問題解答には直接関係ないが後期RNAはみなrif感受性なので、ファージのコードするRNAポリメラーゼはrifampicin感受性か、あるいはサブユニットの場合には rifampicin感受性に関わるユニットと考えられる。 |
|||||||||||||||||||||||||||||||||||||||||||