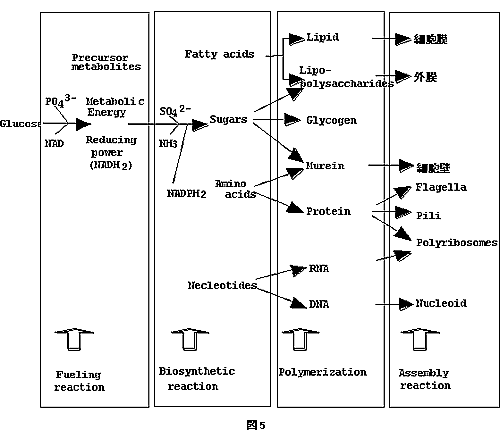
Enterobacteria
は種特有の多彩な発酵経路を持っているので、発酵産物は菌分類の手がかりになる。 Enterobacteria(腸内細菌科)細菌の分類同定にエンテロチューブというものが用いられる。これは沢山の寒天培地を小さなコンパートメントに詰め、針金を中に通したものである。針金の先でコロニーをつつき、針金を引くと、コンパートメントの寒天に 植菌される仕組みになっている。結果は糖を発酵しpHの変化で色が変るか変らないかで見るためコード化出来る。コード番号の表を引くと菌名が出る仕掛になっている。
大腸菌は酸素存在下で aerobic respiration
をし、酸素がなくなると他の電子受容体 NO3 -を利用した anaerobic
respiration をし、それもなくなると、発酵を行う。結核菌はaerobic
respirationしか出来ない。Clostridia(ボツリヌス菌、破傷風菌、ウエルシュ菌)や連鎖球菌は好気性、嫌気性いずれの呼吸も出来ず発酵しか出来ない。
5−2:好気性菌と嫌気性菌
酸素要求性は菌により異なり、分類の鍵になる。次の様に分類する。
- 絶対好気性 (obligate aerobe)
aerobic respirationのみがエネルギー産生系(結核菌)
- 通性嫌気性 (facultative anaerobe )
酸素があってもなくても増殖出来る。これには2種類ある。
大腸菌のように好気性呼吸と嫌気性呼吸、或は発酵が可能なもの。
連鎖球菌のように酸素存下でも発酵しかせず、酸素があっても死なないもの。
- 絶対嫌気性菌 (obligate anaerobe)
酸素存在下で死ぬもの。例えば、Bacteroides、Clostridiumなど。
5−3:真核細胞のエネルギー産生系と共生進化
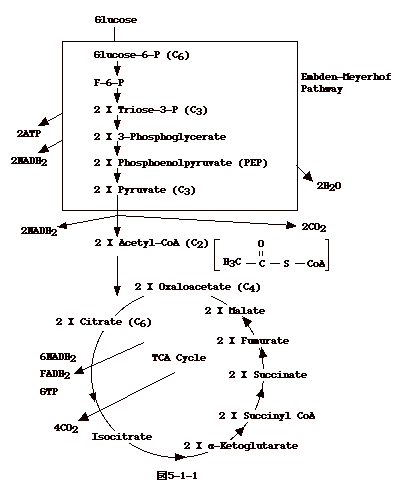
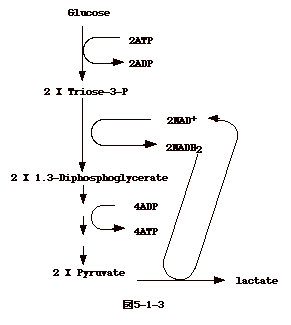
真核細胞のエネルギー産生系はどうなっているのだろうか?真核細胞にも細菌と同じような解糖経路とTCAサイクルがある。
真核細胞ではグルコースから pyruvate への解糖経路(Embden - Meyerhof
経路)は細胞質にある。出来た pyruvate
はミトコンドリアへ運ばれ、TCAサイクルに入る。ここで、6分子のNADH2
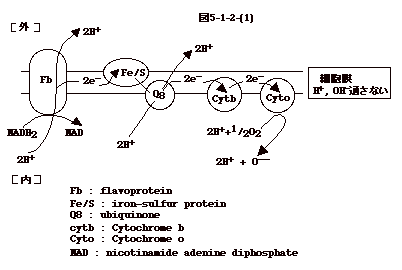
と2分子の FADH2が出来る。 NADH2 と FADH2は、細菌の細胞膜にあるのと同様な機構で
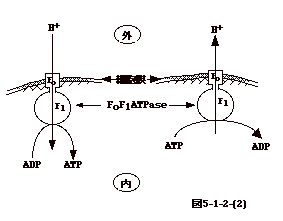
NAD+H2 、FAD+H2に変換され、それぞれの反応で出来た2H+がミトコンドリアの外、つまり細胞質、に放出される。細胞質のH+が、イオン勾配を利用し、ATP
合成酵素であるチャンネルを通りミトコンドリアに入ってくると、ADPは
ATP
に変換される。即ち、基本的にはミトコンドリアの膜が細菌の細胞膜と同じ機能をしてエネルギー産生をしている事になる(図5-3)。
この、H+イオンの膜輸送によるエネルギー産生は、1961年
Peter Mitchellが提唱したもので、それが認められるには 20年以上の月日を必要とした。
ミトコンドリアは細菌と同じ様な環状 DNA
を1コピー持つ。ミトコンドリアが増える時、細菌DNA
と同様な様式で複製される。このDNA は、遺伝子として、NADH
hydrogenase、cytochrome oxidase、cytochrome、ATP合成酵素などエネルギー産生に必要な酵素遺伝子と、22のtRNA
遺伝子、16S及び12S rRNA 遺伝子をコードしている。
上に述べた細菌の機能的、構成的類似性から、ミトコンドリアの起源は細菌細胞の真核細胞内共生にあるのではないか、と云う仮説が出されている(Lynn
Margulis, Symbiosis in Cell Evolution Freeman 1981)。
動物細胞のリボゾームは80S(S値は沈降遠心で物質を分けた時の係数で、概して云えば値の大きい方が重い)で主な
rRNA として 29S、18S、5.8S、5S、の RNA
を持つ。原核細胞のリボゾームは 70Sで、主なRNA は、23S、16S、5SRNA
である。動物細胞のミトコンドリアにある 50ー55S
のリボゾームは機能構造や抗原性において原核細胞リボゾームに似ている。ストレプトマイシンは
70Sリボゾームに特異的に働き細菌の蛋白合成を抑えるが、宿主細胞にも毒性がある。これは、ストレプトマイシンが細菌細胞のリボゾームに似たミトコンドリアのリボゾームにも働く為である。
実際種々の蛋白合成を阻害する抗生物質の作用域はミトコンドリアと細菌で共通している(下表で+は阻害あり、-は阻害なし)